-
LE DANIEN DE VIGNY (sixième partie)
ANALYSE BIBLIOGRAPHIQUE (suite)
Analyse des données concernant les gatéropodes de Vigny (texte de 1990).
Les gastéropodes représentent le groupe de mollusques le mieux représenté, dans les calcaires de Vigny (diversité des espèces et fréquence des individus).
Fréquence relative et mode de vie des espèces.
Les espèces les plus abondantes sont, d'une manière générale, celles qui ont été décrites par d’ Orbigny dans son étude de 1850,
Les diagnoses originales de d’Orbigny toujours très courtes, restent cependant explicites. Les types ont été retrouvés dans les collections de la faculté de Jussieu (coll. Hébert) et du Muséum d'Histoire Naturelle de Paris. Je considère donc ces taxons comme valables (à l'exception de quelques espèces).
Je donnerai ici la fréquence relative des principaux taxons. Cette dernière a été établie comme suit:
- Seront dites abondantes, les espèces que l'on trouve dans tous les lithofaciès (que l'on trouve aisément lors d'une visite sur le gisement).
- Seront dites moyennement abondantes, les espèces que l'on ne trouve que dans des lithofaciès particuliers, mais qu'on est sûr de trouver en une journée de recherche.
- Seront dites fréquentes les espèces que l'on trouve sur 2 ou 3 visites.
- Seront dites moyennent fréquentes celles inféodées à un lithofaciès précis et que l'on trouve en moyenne sur 2 ou 3 visites.
- Les autres taxons, que l'on peut qualifier d'assez rares ou rares ne sont pas pris en compte ici.
Espèces abondantes
- Calliomphallus gravesi: épibionte, herbivore.
- Pleurotomaria penultima: épibionte, herbivore.
- Patella hebertiana: épibionte, herbivore, zone intertidale à infratidale supérieure (adaptation morphologique en fonction du niveau d'énergie du milieu).
- Paleocypraea suecica: épibionte, détritivore (coraux, polypes, etc… ).
- Hipponyx consobrinus: épibionte, fonds essentiellement rocheux, mangeurs de plancton, se fixe sur un support (par l’intermédiaire de son opercule).
- Bivonia vignyensis: endobionte, perforant, sessile, mangeur probable de petites particules végétales.
- Calyptraea supracretacea: épibionte vivant sur des supports divers (autres gastéropodes), herbivore.
Espèces moyennement abondantes.
- Rhinoclavis gea: semi-endobionte vivant dans les boues micritîques crayeuses des Interstices récif aux, détritivoresf brouteurs.
- Rhinoclavis gea var. ornata : semi-endobionte vivant dans les sables profonds, Détrivore ou herbivore.
- Campanilopsis uniplicatum semi-endobionte à épibionte vivant essentiellement dans les masses gravellaires grossières. Herbivore probable.
- Campanilopsis dimorphum semi-endobionte à endobionte vivant dans les dépôts sableux, Détritivore probable.
Espèces fréquentes
- Benoistia pisolithica: épibionte des subtrats très irréguliers.
- Chilodonta geometrica: épibionte; herbivore.
- Jujubinus hannonicus: herbivore…
Espèces moyennement fréquentes
- Campanile maximum : semi-endobionte des substrats sableux. Détrivore probable.
- Hipponyx ornatissimus: épibionte des substrats sableux moyennement indurés.
- Chondrocerithium instabile : endobionte dans les boues crayeuses (faciès mimétique de la craie). Détritivore.
- Charonia rutoti: épibionte, dans tous les lithofaciès. Carnivores: les formes actuelles se nourrissent d’ échinodermes.
- Caricella grandis. épibionte ou semi-endobionte des substrats résistants et irréguliers, carnivore (prédateur probable),
- Scaphella vignyensis: semi-endobionte, environnement gravellaire boueux, carnivore (prédateur probable)*
- Ampullella lavallei: épibionte, substrat gravellaire ou sableux, carnivore, prédateur.
Données écologiques déduites de l' études des gastéropodes.
Les gastéropodes apportent des indications précises pour ce qui concerne l'environnement. Ceci d'autant plus que les espèces sont trouvées sur leur lieu de vie, ou qu'elles n'ont subit qu'un très faible transport.
Les taxons les plus abondants sont tous des épibiontes herbivores. Ceci est évidemment lié à la prolixité des algues calcaires et à l'environnement récifal. Les détritivores sont aussi très nombreux: grosse quantité de matière organique en voie de décomposition, abondance des algues microscopiques…
Certains genres sont représentés par deux espèces proches (ou deux variants d'une même espèce) l'une étant adaptée aux milieux construits indurés et anfractuosaires, l'autre aux sédiments détritiques (grossiers ou fins). C'est le cas pour Campanilopsis, Rhinoclavis, Hipponyx, le couple Scaphella/Caricella…
Les carnivores sont beaucoup moins fréquents que les herbivores et les détritivores. Seule Ampullella peut être considérée comme telle et c'est sans doute à elle qu'il faut attribuer les rares perforations circulaires observées sur les grands cérithes. A Vigny les gastéropodes prédateurs n'ont joués qu'un rôle très secondaire. De fait, la sélection s'opérait par d'autres voies: capture des larves par les polypiers, par exemple…
Données comparatives des faunes de gastéropodes de Vigny avec celles de Maastricht» de Fakse. de Ciply et de Mons.
Sur les 62 genres recensés à Vigny 3 seulement ont un caractère franchement crétacé. Ils appartiennent aux genres Eucycloscala, Odostoma et Tylostoma.
Les gastéropodes communs à Vigny et Mons sont finalement assez peu nombreux: 25 espèces. Soit en pourcentage moyenné: 18% (228 espèces à Mons). Les plus significatives sont: Seila tenuifila, Chondrocerithium instabile; Triforis (E.) montensis; Newtoniella subglabra…
La comparaison avec les faunes de l'Amérique du Nord (Eocène des formations Clairborne par exemple) apporterait sans doute des indications plus signifiantes…
A Vigny les gastéropodes constituent de médiocres indicateurs stratigraphiques. Les résultats sont similaires à ceux résultant de l’étude des lamellibranches. On constate encore une grande ressemblance avec Ciply (38%) ressemblance plus faible avec Fakse (26 %). Et finalement c’est avec Mons que 1’on trouve à Vigny, le moins d'espèces communes (18%)...
Le caractère franchement cénozoïque de l'ensemble des gastéropodes des calcaires de Vigny n’en est pas moins remarquable… Et c’est finalement le seul résultat sur lequel on se doit d’insister à leur propos.
Quelques formes intéressantes …
L’inutile multiplication des taxons ne sert, finalement, ni la compréhension de l’évolution des gastéropodes, ni celle de la synécologie globale… Je me suis borné à signaler les seules espèces vraiment sigificatives : Monodonta (Osilinus) vignyensis, Gibula sarae, Bivonia vigyense, Caricella grandis… en reclassant les taxons décrits par d’Orbigny : Patella hebertiana, Angaria polyphylla, Calliomphalus gravesi, Pseudoliotina danae, Rhinoclavis gea… Ou en rapportant les spécimens à des taxons de Faxe ou de Mons…
L'existence à Vigny d'une vraie patelle (ce n'est ni un Helcion ni un Acmaea…) est très significative. Et les "Helcion" qui connus dans le Maastrichtien, dans le Danien (à Ciply), dans le Montien sont aussi de vraies Patelles…
Les grands Campanilopsis et Campanile semblent être des formes qui caractérisent une période s'étendant du Maastrichtien (Maastricht) au Montien moyen (Mons). II faut toutefois remarquer que l'espèce Campanile maximum est signalée par Hagg (1947) depuis le Cénomanien (z. à mamillatum). Le genre Campanilopsis serait, quant à lui, limité au Paléocène. Au Thanétien les campanilidés sont plus rares. A la fin de l'Eocène (Lutétien-Bartonien) les grands cérithes redeviennent abondants et diversifiés. Ces fluctuations sont, semble-t'il, contrôlées par la température des eaux (au Bartonien les madréporaires redeviennent abondants).
Les cérithidés, marqués par l'apparition des genres Thericium (Globulocerithium) et Benoistia, par la présence de Cerithiella et Cerithiopsis montrent, à Vigny, une faune déjà diversifiée, ce qui accentue le caractère tertiaire de cet ensemble faunistique (sans pour autant augmenter sa ressemblance avec celui de Mons).
On trouve aussi dans les calcaires de Vigny les premiers représentants des genres Zeidora, Monodonta, Gibbula…
- 4 Céphalopodes dont Hercoglossa scanica et Ceratisepia elongata (n. g. et n. sp) sont figurées,

Hercoglossa danica (Schlotheim, 1820). A : Echantillon de la collection Hébert (moulage en cire). E à M: spires juvéniles. O, P et R : gros échantillon (moulage rodorsil). Q, S et T : Aspect de la coquille (moulage rodorsil). Les barres correspondent à 10 mm.
On trouve aussi des Hercoglossa correspondant à la figuration d’ H. schoelleri donnée par Rosenkrantz en 1970. Ceux-ci ont une largeur de spire plus importante, un contour ogival arrondi plus accentué qu’H. danica sensu stricto. Mais iI faut souligner que cette différence n'est pas constante: elle est plus ou moins accentuée d'un individu à l'autre et l’on peut trouver tous les jalons entre la forme danica et la forme schoelleri. De plus certains exemplaires montrent les caractéristiques d’H. danica sur leurs jeunes spires et celles d’H. scheolleri sur les spires adultes ! H. danica et H. schoelleri doivent donc être considérés comme des variants phénotypiques d'une seule et même espèce : Hercoglossa danica (Schloth, 1820). H. danica (schoelleri) a formé de véritables accumulations dans les faciès du premier épisode récifal de Vigny. Elle s'associe alors avec Cimomia heberti.
On trouve aussi de très gros Hercoglossa dont la morphologie s’écarte complètement des taxons signalés précédemment. Ici les individus sont de très grande taille: 550 mm de diamètre! Ils ont une spire surbaissée, peu recouvrante et la suture montre un lobe médiolatéral largement arrondi. Mais surtout le siphon est très oblique au sein des espaces caméraux (v. figure). Cette espèce rare (4 exemplaires) correspond aux figurations de Rosenkrantz (1970) pour sa nouvelle espèce Hercoglossa scanica. Les exemplaires de cette espèce ont été trouvés en un seul point du gisement de Vigny au milieu d'une multitude d'arches (Barbatia gravesi) ayant leurs deux valves en connection…

Hercoglossa scanica (Rosenkrantz, 1970). A : Representation schématique de l'échantillon le plus complet (perdu lors d'un déménagement) Barre = 100 mm. B à F : Chambres d'habitation. Barres = 100 mm. G et H : Moules internes dissociés des locula. Barres = 70 mm. I : Moule interne d'une chambre d'habitation montrant de nombreux et très grands serpulidés (empreintes externes). Barre = 10 mm. J à L : Moule interne d'une locula montrant la disposition très oblique du siphon. Barres = 10 mm. M à O : moules naturels du siphon soulignant sa taille relativement importante et l'obliquité de ses attaches. M et O : attaches adorales. N : attaches adapicales. Barres = 10 mm.
Ajoutons une autre espèce d’Hercoglossa, probablement nouvelle, caractérisée par une spire surbaissée, et une selle latérale très marquée et profonde dont le siphon semble moins oblique que sur H. scanica (espèce auquelle j’avais d’abord rapporté cet échantillon).
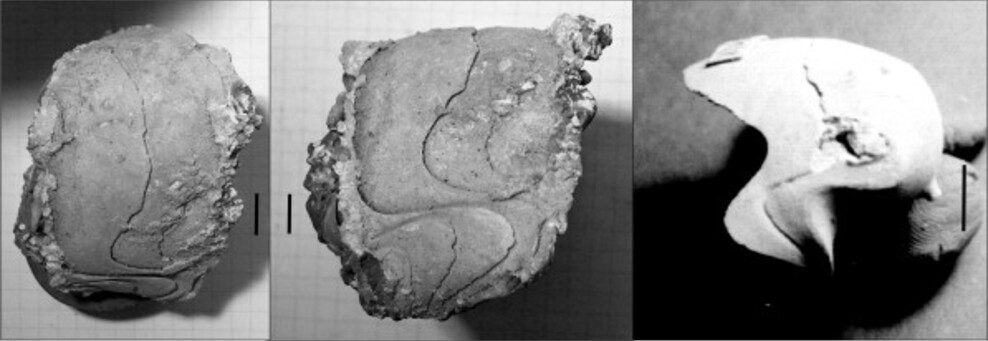
Hercoglossa sp. Spire surbaissée, selle profonde. Les barres correspondent à 10 mm.
Cimomia herberti (d’Orbigny, 1848) est, avec Hercoglossa danica très abondant à Vigny. Cette espèce, qui peut atteindre également une taille relativement importante (180 mm de diamètre), se caractérise par son galbe subglobuleux, sa section spirale réniforme, son épaulement ombilical épais, par une suture dessinant une selle périomblicale et par son ombilic très étroit.
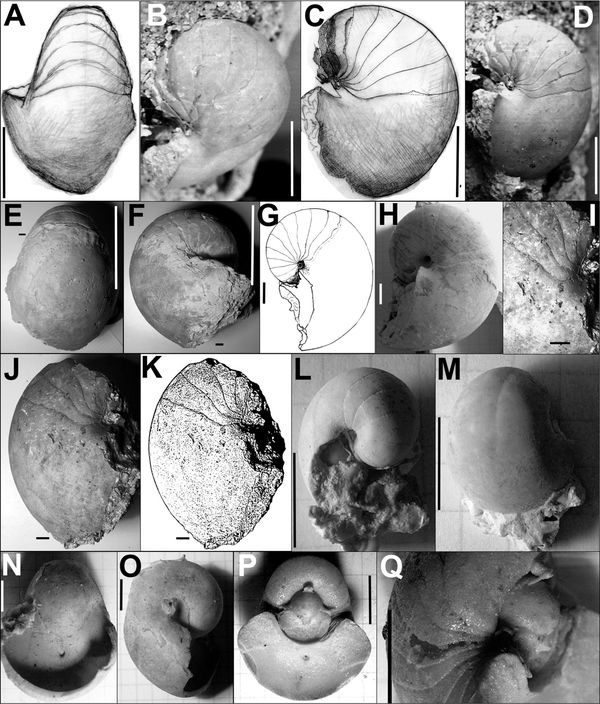
Cimomia herberti (d’Orbigny, 1848) A à M : série d'échantillons conservant leur chambre d'habitation. Les empreintes musculaires sont obscures, peu visibles.. A à F : forme adulte. Barres = 100 mm. Noter la présence d'une sele latérale obtuse. G et H : adulte de petite taille. Barres = 10 mm. J et K Adulte. Barres = 10 mm. L et M : forme juvénile. Notre les lignes septales rectilignes, la spire relativement haute et l'ombilic assez large. Barres = 10 mm. N et O : moules rodorsil. Aspect original de la coquille. Cimomia heberti montrait un cal omblical épais et son siphon était étroit. P et Q : Moule interne montrant l'ornementation transversale affectant la coquille juvénile. Barres = 10 mm.
Quelques fragments d'un nautile plus régulièrement globuleux, quasisphérique et sans selle péri-ombilicale (la ligne suturale est droite) sont rapportés à Eutrephoceras sp. II s'agit peut être d' E. bellerophon, Lundgren 1867 qui est cité du Dano-Montien par Furon & Soyer (1947, p, 16).

Eutrephoceras cf. bellerophon, Lundgren 1867. A à E : Echantillon avec chambre d'habitation (A à C). Les empreintes musculaires sont profondes. Les sutures sont lègèrement sinueuses mais ne dessinent pas de selle peri-ombilicale. Ombilic relativement large. Coquille globuleuse. Barres = 10 mm. F : Coquille partiellement épigenisée montrant un large siphon sub-central. Barre = 10 mm. G : Moule interne. Barre = 10 mm.
Ceratisepia elongata, n.gen, n. sp..
Une place à part doit être faite à Ceratisepia elongata, n.gen., n.sp., d’ une part parcequ’ il s’agit ici d’un sépidé (Ordre regroupant les seiches et les spirules actuelles) et non d'un nautile; d'autre part parce que sa présence à Vigny, sans être directement un argument stratigraphique, n'en est pas moins le témoin significatif de la période de transition que représente le Danien.
En effet C. elongata est une forme archaïque, une forme ancestrale pour les Sepiidés. Ceux-ci n'existent pas dans le Crétacé (Groenlandibelus, Jeletzky, du Maestrichtien groenlandais, est très probablement un Belemnitidé Diplobeliné).
On trouve dans le Montien de Mons deux Sépidés assez proche de Ceratisepia mais qui s'en distinguent par la présence d'un capuchon rostral bien développé. Ils ont été rapportés au genre Beloptera par Briart et Cornet en 1877, Ils doivent être reversés dans des genres différents genres conduisant à Beloptera d'une part et à Belosepia d'autre part. Par rapport à ceux-ci Ceratisepia montre des structures nettement moins différenciées. Ceci n'est évidemment pas surprenant si l'on considère les assises de Vigny (et le Danien dans son ensemble) comme une entité stratigraphique antérieure au Montien (ce qui reste discuté).

Ceratisepia elongata (Meyer, 1987) A et B : moule interne de la portion apicale du phragmocône. Barres = 10 mm.C : moule interne des deux dernières loges adorales et du siphon. Barre = 10 mm. D et E : aspect du capuchon rostral et du phragmocône. Moulage en rodorsil. Cap. = capuchon rostral. Barre = 10 mm. F et G : Capuchon rostral en gros plan. Moulage rodorsil. av = apophyse ventrale. cap = capuchon rostral. Barre = 10 mm. J et K : Moule interne de deux loges et d'une portion de la cavité siphonale. Barres = 10 mm. L : Vue ventro-apicale du moule interne d'une coquille juvénile. Barre = 10 mm. M : Moule interne de la partie apicale du phragmocône montrant la protoconque. Barre = 5 mm.
Implications et données déduites à partir de l'étude des céphalopodes de Vigny.
A Vigny, les céphalopodes ne sont fréquents qu'à la base des formations récifales, c'est-à-dire dans les calcaires bioclastiques inférieurs, dans les assises du premier épisode récifal. Mais on retrouve les mêmes espèces, beaucoup plus dispersées, et généralement brisées, dans toutes les autres assises.
La relative abondance des nautiles àla base des formations calcaires de Vigny s'explique par les modalités de l'édification récifale. Durant le premier épisode récifal les pâtés récifaux sont d'abord séparés par de larges passes s'ouvrant sur la haute mer. Les nautiles transportés par flottaison franchissent facilement l'alignement des masses récifales et viennent s'échouer le long des escarpements crayeux alors partiellement exondés (falaises). Durant la seconde phase récifale, les masses construites forment une barrière plus continue et empêchent les coquilles transportées par flottaison de se déposer dans les zones d'arrière récif, dont les sédiments sont, pour ce niveau, seuls observables aujourd'hui.
La préservation des nautiles, qui conservent la plupart du temps leur chambre d'habitation, et de Ceratisepia (seul sépiidé fossile connu avec ses structures phragmoconiques bien conservées) dans un sédiment s'étant déposé en mileu agité (pendages anormaux traduisant l'existence de chenaux, de rides…) reste surprenante. On peut l'expliquer comme suit. Les premiers dépôts daniens se sont réalisés au pied d'escarpements crayeux très irréguliers partiellement exondés. Les courants ont entraînés les coquilles des céphalopodes dans des zones abritées où les boues crayeuses issus de l'érosion littorale s'accumulaient en abondance. La proximité du rivage et la nature du climat favorisait aussi l'apport d'argiles.
Le taux de sédimentation élevé a permis un enfouissement rapide des coquilles, en les préservant d'une destruction mécanique immédiate. Il est problable que la teneur relativement importante en terrigènes des sédiments côtiers, teneur accentuée dans les zones de faible dynamique, a préservé les tests aragonitiques de ces organismes, d'une dissolution rapide. Les tests ont d'abord été encroûtés d’aragonite palissadique (aujourd'hui transformée en une croûte microsparitique).
La diagenèse a, ensuite, été très lente. L'aragonite des tests n'a été mobilisée qu'après lithification complète de l'orthochème, préservant ainsi les vides coquilliers d'un remplissage sédimentaire (porositéde moule). Les ciments ont ensuite subit une dernière phase de recristallisation après exondation des sédiments (sparite drusique).
La prolifération des nautilidés à la base du Paléocène est un fait évident et significatif. Les nautiles, animaux vagiles, donc occupant une vaste aire géographique, impliquent aussi des eaux de surface relativement chaudes. Cette condition, comme d'ailleurs l'implantation d'édifices récifaux à de hautes latitudes, est liée au contexte paléogéographique de l'époque. L'élargissement de l'Atlantique Nord bordédans sa partie septentrionale par les hauts fonds qui relient la Scandinavie au Groenland, le rétrécissement du domaine mésogéen ont vraisemblablement entraîné une modification du parcours des courants équatoriaux. Ceux-ci furent déviés vers l'Europe. Ils ont favorisés la prolifération des organismes de mer chaude et donc l'implantation d'édifices récifaux.
L'ouverture de l'Atlantique Nord génère au début du tertiaire l'arrivée de nouveaux organismes, la compétition interspécifique sans pour autant entraîner un refroidissement des eaux (période sub-sahélienne ?). Durant une courte période, l'irruption des eaux arctiques se réalise au travers de l’échange des eaux de surface, pourvoyeuses des formes nouvelles planctoniques et vagiles (foraminifères planctoniques, et peut-être, aussi, des sépiidés). Cet événement suffit pour rompre la chaîne alimentaire qui conduit aux ammonites et aux bélemnites et provoque leur disparition. Disparition d’autant plus rapide que ces derniers relictuels et hyper spécialisé ont amorcé leur déclin dès le Sénonien… D’ailleurs leur disparition est quasi-effective durant le Maestrichtien supérieur.
Rapidement, les nautiles occupent, pour la courte durée qui précède l'arrivée en masse des eaux froides arctiques (s'amorçant au cours du Montien et atteignant son maximum au Thanétien), la niche écologique laissée vacante par les ammonites.
Au Danien, les courants chauds déviés vers l'Est par les hauts fonds nordiques (large plateforme joignant la Scandinavie, le Groenland et l'Amérique du Nord) favorisent l'implantation très septentrionale des complexes récifaux… Simultanément à la prolifération des nautiles.
Au Montien la séparation Scandinavie-Groenland s'accentue. Sous l'influence marquée des eaux froides nordiques les récifs disparaissent. Mais les eaux de surface toujours influencées par les courants équatoriaux restent chaudes: les nautiles continuent de proliférer avec des espèces différentes.
Plus tard, la séparation complète de la Scandinavie d'avec le Groenland entraîne une profonde modification du régime des courants (issus des zones nord-orientales). Corrélativement on constate un refroidissement général de la région Nord-Atlantique (phase où l'eau est piégée sous forme de glace -période glaciaire ?). Survient alors un retrait général de la mer (Thanétien inférieur). L'arrivée des eaux thanétiennes par le Nord-Est (fonte partielle des calottes glaciaires ?), est marquée par un réchauffement relatif (algues dasycladales abondantes). Mais les nautiles, soumis aux courants chauds venants du Sud (paléo-gulf-stream), sont alors relictuels et représentés de façon plus marginale.
...
- 17 Ostracodes, 3 figurés: Curfsina geleenensis, Paleomonsmirabilia geulensis, Limburgina bilamellosa bilamellosa.
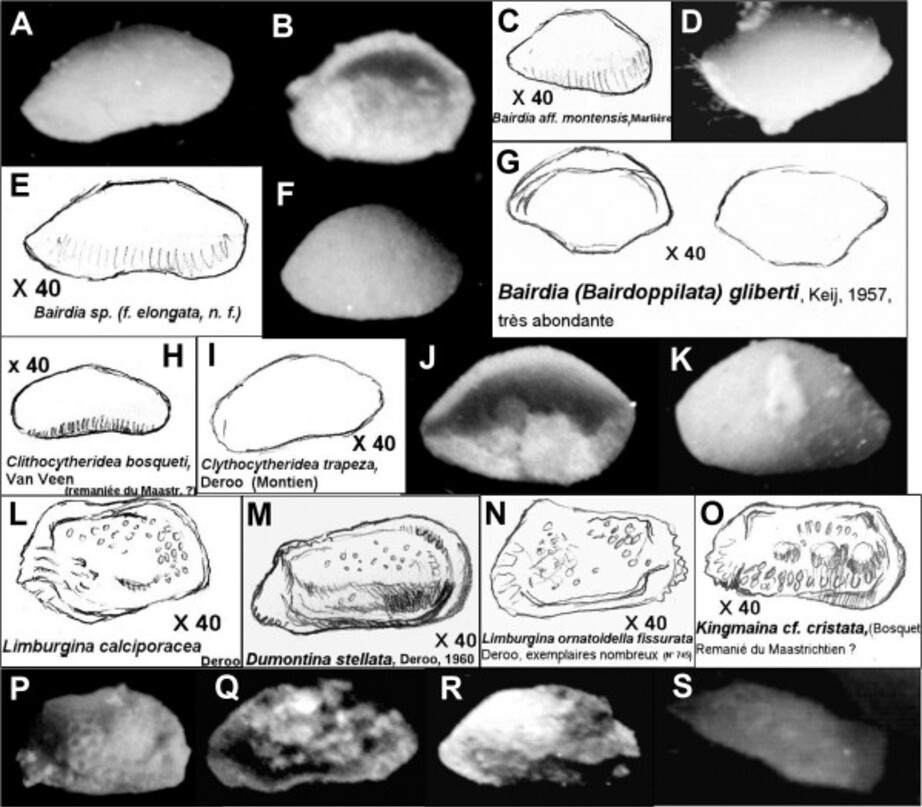
Je donne ici quelques schémas originaux et photos d'ostracodes. Les déterminations sont sujettes à caution et ne sont indiquées que pour information. Les calcarènites de Vigny recelaient une quantité remarquable de ces petits organismes. Leur étude complète reste à réaliser...
OSTRACODES (texte de 1990)
1 - Références bibliographiques.
Je me suis ici servi des travaux de Grekoff (1956), Marlière (1958), Damotte & Feugueur (1963), Damotte (1964, 1971, 1986)) et Deroo (1966),
2 - Les espèces des calcaires de Vigny.
Bairdia (Bairdoppilata) gliberti, Keij, 1957
Bairdia montensis Marlière, 1958
Bairdia sp,
Clithocytheridea trapeza, Deroo, 1965
Clithocytheridea bosqueti, Van Veen, 1936
Cytherella sp.,
Paleomonsmirabilia geulensis, Deroo, 1965
Paracytheretta aff. northensis, Deroo, 1966
Eucytherura simplex, Van Veen, 1935
Curfsina geleenensis, Deroo, 1966
Dumontina stellata, Deroo, 1965
Limburgina ornatoidella fissurata, Deroo, 1965
Limburgina bilamellosa bilamellosa, (Marlière, 1958)
Limburgina calciporacea, Deroo, 1965
Limburgina subornatella, Deroo, 1965
Limburgina longiporacea, Deroo, 1965
Amphicytherura sp.
3 - Données déduites.
Le matériel nouveau a permis la reconnaissance de quelques espèces supplémentaires. Aux 9 espèces signalées par Damotte & Feugueur à Vigny (1963) s'ajoutent 8 autres espèces recueillies dans les sables de Vigny. Mais il est très probable que de nombreux autres taxons existent à Vigny. L'étude approfondie des faunes d'ostracodes reste à faire.
Disons le tout net: aucun des genres reconnus à Vigny n'est, en soi, caractéristique du Tertiaire. Tous existent dans les assises sénoniennes et maastri chtiennes. Les espèces qui caractérisent la base du Tertiaire et plus précisément le Montien, Cytherella lagenalis, Krithe montensis existent dans le Montien de Montainville, de Mons et du Limbourg mais sont absentes à Vigny. La présence de Bairdia montensis n'est, semble t’il, guère signifiante car des formes très proches existent dans la craie campanienne: B. cuvillieri, Damotte, 1971 par exemple.
Trois espèces sont fréquentes: Bairdia (Bairdoppilata) gliberti, Limburgina bilamellosa bilamellosa et Limburgina ornatoidella fissurata. Ce sont précisément celles-là qui ont permis la définition des assises à Cytherelloidea par Marlière en 1958 et qui correspondent très précisément au tuffeau de Ciply que l'on considère, depuis Rasmussen (1964), comme Danien. L. ornatoidella fissurata correspond, à Vigny, aux figures 743-746 de Deroo. Elle est par contre très différente de la forme géante représentée par le même auteur dans les figures 734-736.
La présence dans notre gisement des espèces suivantes connues dans le Danien du Limbourg: Curfisina geleenensis (également connu à Fakse), Limburgina calciporacea, Limburgina subornatella, Limburgina longiporacea pourraient confirmer la position stratigraphique des assises de Vigny considérées comme légèrement inférieures à celles de Ciply. On notera aussi à Vigny l'existence de quelques espèces maastrichtiennes, représentées par de rares échantillons usés et semble-t’il remaniés.
Les études références portent sur les Ostracodes de la région de Mons (Marlière 1958) et sur ceux du Limbourg (Deroo, 1966). Rien, ou très peu de chose n’est connu (ce texte date de 1990) pour ce qui concerne les ostracodes du Danien de Fakse. Or dans les deux régions où les ostracodes ont été suffisamment étudiés, on note une lacune du Danien inférieur et du Danien moyen. En conséquence l'extension verticale des espèces citées est tronquée vers le bas… D'autre part des formes telles que D. stellata, et L. bilamellosa bilamellosa couvrent AU HOINS le sommet du Danien (tuffeau de Ciply) et la base du Montien…
Les trois espèces significatives sont C. geleenensis, P. geulensis et L. bilamellosa bilamellosa. Ces taxons connus du Limbourg, de Ciply et de Fakse ont une extension stratigraphique limitée au SEUL Danien moyen/supérieur !
Les ostracodes pourraient donc confirmer les résultats obtenus à partir de l’étude des foraminifères, des brachiopodes et des céphalopodes. Les assises récifales de Vigny sont à rapporter au Paléocène inférieur et plus précisément à la limite Danien moyen/danien supérieur. Ils sont légèrement antérieurs aux calcaires de Ciply. . .
Les formes trouvées à Vigny traduisent la proximité du littoral (Bairdia), une salinité normale et une profondeur comprise entre 20 et 50 m.
CIRRIPEDES (texte de 1990)
Les cirripèdes acrothoraciques qui perforent les tests des mollusques et des coraux ont certainement joués, au même titre que les cliones, le rôle de pourvoyeurs en micrite. De ce point de vue, ces organismes, souvent passés sous silence, doivent être considérés comme un élément important dans l'écosystème récifal. Mais leurs perforations atypiques n'en autorisent aucune détermination.
-
Commentaires
|
|
|
|